Что отвечает за дыхание в головном мозге
Величина легочной вентиляции обусловлена частотой и глубиной дыхательных движений (дыхательного ритма), происхождение которых связано с функцией дыхательного центра центральной нервной системой. Под дыхательным центром понимают ограниченный участок ЦНС, где происходит формирование дыхательного импульса, вызывающего координированную деятельность дыхательных мышц, обеспечивающих для организма необходимую величину газообмена в легких. В центральной нервной системе местом генерации дыхательного ритма, вызывающего ритмические сокращения дыхательных мышц при вдохе и выдохе, является продолговатый мозг, в котором расположен дыхательный центр. Дыхательный центр состоит из нервных клеток (дыхательных нейронов), для которых характерна периодическая электрическая активность в одну из фаз дыхания. Нейроны дыхательного центра локализованы двусторонне в продолговатом мозге в виде двух вытянутых столбов вблизи obex — точки, где центральный канал спинного мозга впадает в четвертый желудочек. Эти два образования дыхательных нейронов в соответствии с их положением относительно дорсальной и вентральной поверхности продолговатого мозга обозначают как дорсальная и вентральная дыхательные группы (рис. 10.21).
Дорсальная дыхательная группа нейронов образует вентролатеральную часть ядра одиночного тракта. Дыхательные нейроны вентральной дыхательной группы расположены в области n. ambiguus каудальнее уровня obex, n. retroambigualis непосредственно ростральнее obex и представлены комплексом Бетзингера, который находится непосредственно вблизи n. retrofacialis вентролатеральных отделов продолговатого мозга. В состав дыхательного центра входят нейроны двигательных ядер черепно-мозговых нервов (обоюдное ядро, ядро подъязычного нерва), которые иннервируют мышцы гортани и глотки.
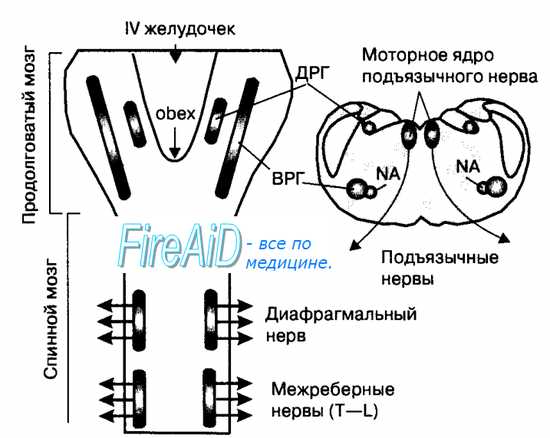 Рис. 10.21. Схема положения нейронов дорсальной (ДРГ) и вентральной дыхательных групп (ВДГ) дыхательного центра продолговатого мозга. Слева — проекция отделов дыхательного центра на дорсальную поверхность продолговатого мозга. Справа — расположение отделов дыхательного центра на поперечном плане продолговатого мозга. Диафрагмальные мотонейроны и мотонейроны межреберных мышц являются «общим конечным путем» моторных команд дыхательного центра, обусловливающих сокращение дыхательных мышц и вентиляцию легких. NA — n. ambiguus.
Рис. 10.21. Схема положения нейронов дорсальной (ДРГ) и вентральной дыхательных групп (ВДГ) дыхательного центра продолговатого мозга. Слева — проекция отделов дыхательного центра на дорсальную поверхность продолговатого мозга. Справа — расположение отделов дыхательного центра на поперечном плане продолговатого мозга. Диафрагмальные мотонейроны и мотонейроны межреберных мышц являются «общим конечным путем» моторных команд дыхательного центра, обусловливающих сокращение дыхательных мышц и вентиляцию легких. NA — n. ambiguus.
Основным критерием классификации нейронов дыхательного центра является фаза дыхательного цикла, в которую они активны, т. е. инспирация или экспирация. По этому критерию дыхательные нейроны подразделяют на инспираторные и экспираторные. Дорсальная дыхательная группа состоит полностью из инспираторных нейронов. Вентральная дыхательная группа образована инспираторными и экспираторными нейронами, а комплекс Бетзингера образуют только экспираторные нейроны. По паттерну электрической активности нейронов в пределах фаз дыхательного цикла инспираторные и экспираторные нейроны подразделяют на нейроны с нарастающим, постоянным или декрементным типом активности (рис. 10.22).
По проекции аксонов дыхательные нейроны разделяют на премоторные или бульбоспинальные нейроны и проприобульбарные. Аксоны премоторных дыхательных нейронов переходят на противоположную сторону продолговатого мозга, а затем направляются к мотонейронам спинного мозга. Функция инспираторных премоторных дыхательных нейронов заключается в управлении электрической активностью инспираторных мотонейронов диафрагмы и наружных межреберных мышц во время их сокращения при вдохе. В обычных условиях экспирация осуществляется пассивно, поэтому функция экспираторных премоторных дыхательных нейронов реализуется только при увеличении глубины дыхательных движений. Премоторные нейроны комплекса Бетзингера выполняют уникальную функцию — они тормозят все типы инспираторных нейронов дыхательного центра и диа-фрагмальные мотонейроны. Поэтому их аксоны распределяются билатерально, т. е. направляются к соответствующим нейронам, расположенным как ипсилатерально, так и контралатерально.
Рис. 10.22. Образцы электрической активности основных типов нейронов дыхательного центра. Наиболее общая классификация типов дыхательных нейронов основана на связи их электрической активности с фазами вдоха и выдоха дыхательного цикла и паттерном разряда потенциалов действия. По этому критерию классифицируют: полные инспираторные с нарастающей активностью (1), экспираторные нейроны с нарастающей активностью (2), ранние инспираторные нейроны с декрементной активностью (3), поздние инспираторные нейроны с нарастающей активностью (4), экспираторные или постинспираторные нейроны с декрементной активностью (5), полные инспираторные нейроны с постоянной активностью (6).
Аксоны проприобульбарных дыхательных нейронов (ранние инспираторные, постинспираторные, поздние инспираторные, экспираторные нейроны комплекса Бетзингера) оканчиваются на мембране нейронов самого дыхательного центра, расположенных в вентральной дыхательной группе. Функция большинства проприробульбарных нейронов заключается в генерации дыхательного ритма.
Что отвечает за дыхание в головном мозге
3.1 Нервная регуляция дыхания
Дыхательный центр представляет собой совокупность нейронов продолговатого мозга, обладающих ритмической активностью и определяющих ритм дыхательных движений. Бульбарный дыхательный центр выполняет две основные функции:
1) регуляцию двигательной активности дыхательных мышц (двигательная функция);
2) гомеостатическую, связанную с изменением характера дыхания при сдвигах газового состава и кислотно-основного равновесия в крови и тканях.
Двигательная функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна (длительности вдоха, выдоха, величины дыхательного объема).
Нейроны дыхательного центра расположены в дорсомедиальной и вентролатеральной областях продолговатого мозга, образуя так называемую дорсальную и вентральную дыхательные группы. В указанных дыхательных группах расположены следующие виды нейронов:
1) ранние инспираторные, максимальная частота разряда которых приходится на начало инспирации;
2) поздние инспираторные нейроны, максимальная частота разряда – в конце инспирации;
3) полные инспираторные нейроны, характеризующиеся постоянной активностью в течение фазы вдоха;
4) постинспираторные нейроны, максимальный разряд которых обнаруживается в течение выдоха;
5)экспираторные нейроны, активность которых возрастает во второй части выдоха;
6) преинспираторные нейроны, максимальный пик активности проявляют перед началом вдоха.
В структурах бульбарного дыхательного центра различают так называемые респираторно-связанные нейроны, активность которых совпадает с ритмом дыхания, но они не иннервируют дыхательные мышцы, а обеспечивают иннервацию верхних дыхательных путей.
В соответствии с локализацией нейронов бульбарного дыхательного центра, различают дорсальную дыхательную группу (ДДГ) и вентральную дыхательную группу (ВДГ). Нейроны дорсальной дыхательной группы получают афферентные сигналы от легочных рецепторов растяжения по волокнам n. Vagus. Только часть инспираторных нейронов дорсальной группы дыхательного центра связана аксонами с дыхательными мотонейронами спинного мозга, преимущественно с контрлатеральной стороной.
Вентральная дыхательная группа расположена латеральнее обоюдного ядра продолговатого мозга, подразделяется на ростральную и каудальную части. Причем, ростральная часть вентральной дыхательной группы представлена ранними, поздними, полными инспираторными и постинспираторными нейронами.
Дорсальная и вентральная группы нейронов в правой и левой половинах продолговотого мозга взаимосвязаны как в пределах одной половины, так и с нейронами противоположной стороны. В синхронизации деятельности контрлатеральных нейронов бульбарного дыхательного центра участвуют проприобульбарные нейроны и экспираторные нейроны комплекса Бетцингера.
Таким образом, нейроны бульбарного дыхательного центра в зависимости от их значимости в регуляции внешнего дыхания разделяют на три группы:
1) нейроны, иннервирующие мышцы верхних дыхательных путей и регулирующие поток воздуха в дыхательных путях;
2) нейроны, синаптически связанные с мотонейронами спинного мозга и регулирующие активность мышц вдоха и выдоха;
3) проприобульбарные нейроны, участвующие в генерации дыхательного ритма, аксоны которых обеспечивают связь только с нейронами продолговатого мозга.
Подобно многим физиологическим системам контроля, система управления дыханием организована как контур отрицательной обратной связи.
Афферентация с различных рецепторных зон интегрируется в бульбарном дыхательном центре. Последний, в свою очередь, генерирует импульсацию к мотонейронам спинального отдела дыхательного центра, регулирующего сократительную активность дыхательной мускулатуры.
Важная роль в регуляции внешнего дыхания отводится центрам варолиева моста, в частности, пневмотаксическому центру. Последний включает медиальное, парабрахиальное ядро и ядро Келликера. В парабрахиальном ядре находятся преимущественно инспираторные, экспираторные и фазопереходные нейроны. Ядро Келликера содержит инспираторные нейроны.
Дыхательные нейроны моста участвуют в механизмах смены фаз дыхания, регулируют величину дыхательного объема.
Непосредственными регуляторами сократительной способности дыхательных мышц являются спинальные мотонейроны, получающие информацию по нисходящим ретикулоспинальным путям от бульбарного дыхательного центра.
Как известно, нейроны диафрагмального нерва расположены узким столбом в медиальной части вентральных рогов от СIII до CV. Подавляющее количество волокон диафрагмального нерва являются аксонами α-мотонейронов, а меньшая часть представлена афферентными волокнами мышечных и сухожильных веретен диафрагмы, а также рецепторов плевры, брюшины и свободных нервных окончаний самой диафрагмы.
Мотонейроны, иннервирующие межреберные мышцы, расположены в передних рогах спинного мозга на уровне TIV-TX, из них часть нейронов регулирует сокращения межреберных мышц, а другая часть – их позно-тоническую активность.
Обращает на себя внимание тот факт, что активность спинальных мотонейронов, обеспечивающих регуляцию двигательной активности межреберных мышц и диафрагмы, в свою очередь, находится под контролем инспираторных нейронов спинного мозга, расположенных на уровне СI-CII вблизи латерального края промежуточной зоны серого вещества.
В обеспечении дыхания, особенно в условиях патологии, участвуют мышцы брюшной стенки, получающие иннервацию от мотонейронов спинного мозга на уровне TIV-LIII.
Двум фазам внешнего дыхания (вдоху и выдоху) соответствуют три фазы активности бульбарного дыхательного центра: инспирация, пассивная контролируемая экспирация и активная экспирация. Во время фазы инспирации диафрагма и наружные межреберные мышцы увеличивают силу сокращения, активируются мышцы гортани, расширяется голосовая щель, снижается сопротивление потоку воздуха. В постинспираторную фазу дыхания происходит медленное расслабление диафрагмы, сокращение мышц гортани, выход воздуха в окружающую среду.
В фазе экспирации – экспираторный поток усиливается за счет сокращения внутренних межреберных мышц и мышц брюшной стенки.
Рефлекторная регуляция дыхания обеспечивается за счет афферентной импульсации в бульбарный дыхательный центр с различных рецепторных зон. Мощной рефлексогенной зоной является слизистая оболочка полости носа, где расположены различные типы механорецепторов, в том числе ирритантные, растяжения, а также болевой чувствительности, обоняния.
Возбуждение этих рецепторов возникает в момент каждого вдоха и приводит к формированию потока афферентной импульсации в ретикулярную формацию ствола мозга с последущей активацией бульбарного дыхательного центра, сосудодвигательного центра, гипоталамических и корковых структур мозга.
Раздражение ирритантных рецепторов слизистой оболочки носа приводит к рефлекторному сужению бронхов, голосовой щели, остановке дыхания на выдохе, развитию брадикардии, а в ряде случаев прекращению сердечных сокращений и другим изменениям (тормозной тригемино-вагусный рефлекс Кречмера ).
Слизистая трахеи и бронхов является слабой рефлексогенной зоной. В стенке крупных внелегочных бронхов и трахеи имеются высокопороговые, низкочувствительные медленноадаптирующиеся, быстроадаптирующиеся и промежуточные механорецепторы, в норме их роль в регуляции дыхания минимальна.
Чувствительность этих рецепторов возрастает при развитии воспалительного процесса в бронхолегочной системе инфекционной или аллергической природы, когда освобождаются медиаторы воспаления и аллергии: гистамин, кинины, лейкотриены, простагландины и др.. Возбудимость рецепторов трахеи и бронхов возрастает и в случае застойных явлений в малом кругу кровообращения, когда прежние объемы воздуха сильно растягивают стенки воздухоносных путей. Афферентация с рецепторов трахеи и бронхов направляется в бульбарный дыхательный центр по чувствительным волокнам n. Vagus, модулируя глубину и частоту дыхательных движений.
Мощной рефлексогенной зоной является паренхима легких, обеспечивающая не только альвеолярное дыхание, но и рефлекторную регуляцию внешнего дыхания.
Основные типы легочных вагусных афферентов включают: медленноадаптирующиеся рецепторы растяжения альвеол, быстроадаптирующиеся рецепторы, С-волокна.
Многочисленные быстроадаптирующиеся рецепторы (БАР) находятся в эпителии внутрилегочных бронхов и бронхиол. Эти рецепторы наиболее чувствительны к следующим типам раздражителей: ирритантным воздействиям, повреждению паренхимы и механическому раздражению дыхательных путей. Возбуждение БАР возникает также при глубоком дыхании, легочной эмболии и капиллярной гипертензии. Афферентация с этих рецепторов распространяется по чувствительным маломиелинизированным волокнам n. Vagus в ретикулярную формацию ствола мозга и бульбарный дыхательный центр, вызывая бронхоконстрикцию, тахипноэ, развитие кашля и тахикардии. Возбуждение этих рецепторов может быть клинически значимым в патогенезе бронхиальной астмы и нарушениях реактивности дыхательных путей.
По данным ряда авторов в паренхиме легких выделяют и БАР рецепторы спадения, реагирующие на спадение альвеол под воздействием внутрилегочных и внелегочных факторов. Афферентация с этих рецепторов поступает в бульбарный дыхательный центр по маломиелинизированным волокнам n. Vagus и обеспечивает развитие тахипноэ.
Медленноадаптирующиеся рецепторы растяжения – важная группа механорецепторов c вагусной афферентацией, расположенных в гладких мышцах воздухоносных путей. Частота импульсов с этих рецепторов возрастает по мере растяжения альвеол вдыхаемым воздухом и распространяется по толстым миелинизированным α-волокнам n. Vagus в бульбарный дыхательный отдел, обеспечивая формирование рефлекса Геринга-Брейера. Последний контролирует частоту и глубину дыхания, имеет физиологическое значение при дыхательных объемах превышающих 1 л (у взрослых при физической нагрузке). Рефлекс Геринга-Брейера более важен для регуляции дыхательного акта у новорожденных, а также в условиях патологии как один из механизмов реализации инспираторной, экспираторной и смешанной одышек.
Третьей группой легочных механорецепторов являются С-волокна – тонкие миелинизированные вагусные афференты. С – волокна оканчиваются в паренхиме легких, в бронхах и кровеносных сосудах, активируются экзогенными раздражителями и медиаторами альтерации. Активация С-волокон приводит к тахипноэ, брадикардии, гиперсекреции слизи. В состав С-волокон входят J-рецепторы, расположенные в альвеолярных перегородках в контакте с капиллярами (юкстакапиллярные рецепторы), чувствительные к интерстициальному отеку, легочной венозной гипертензии, микроэмболии, раздражающим газам и ингаляционным наркотическим веществам. Активация J-рецепторов вызывает закрытие гортани и апноэ, за которыми следует частое поверхностное дыхание, гипотензия и брадикардия.
Важная роль в рефлекторной регуляции дыхания отводится проприорецепторам суставов грудной клетки, межреберных мышц, диафрагмы, сухожильным рецепторам. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которая через α-мотонейроны повышает активность α-мотонейронов и дозирует таким образом мышечное усилие.
В регуляции активности бульбарного дыхательного центра и внешнего дыхания принимает участие и афферентация с висцеральных рецепторов и рецепторов кожи, о чем свидетельствует развитие гипервентиляции легких при болевом и термическом раздражении.
3.2. Механизмы гуморальной регуляции дыхания
Важная роль в регуляции дыхания отводится хеморецепторам.
Изменения газового состава крови (РаО2, РаСО2) влияют на активность дыхательного центра путем возбуждения хеморецепторов каротидных и аортальных телец (периферические рецепторы), а также хеморецепторов вентральной зоны продолговатого мозга и дорсального дыхательного ядра (центральные рецепторы). Периферические хеморецепторы (рис.5) обеспечивают регуляцию частоты дыхательных движений. Адекватным раздражителем для них является уменьшение РО2 артериальной крови, в меньшей степени – увеличение РСО2 и снижение рН. Периферические хеморецепторы расположены у бифуркации общих сонных артерий на внутреннюю и наружнюю. Несмотря на свой миниатюрный размер, каротидные тельца интенсивно кровоснабжаются (1,4-2 л/мин на 100 г ткани). Этот орган особенно чувствителен к колебаниям кислорода в артериальной крови. При Ра О2 в пределах 60-80 мм рт. ст. наблюдается слабое усиление вентиляции, при Ра О2 ниже 50 мм рт. ст. возникает выраженная гипервентиляция легких. Ра СО2 и рН крови потенцируют эффекты гипоксемии на артериальные хеморецепторы и не являются адекватными раздражителями для этих рецепторов. После двустороннего удаления каротидных телец гипоксический вентиляторный ответ у человека исчезает. При отсутствии хеморецепторной стимуляции, например, при глубокой гипокапнии, повреждении синокаротидной зоны (опухоли, коллагенозы, травмы) ритмогенез дыхания снижается и полностью прекращается.
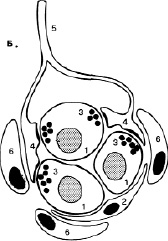
Рис. 5. Каротидное тельце: 1-хеморецепторные клетки; 2-поддерживающие клетки; 3-синаптические пузырьки; 4-чувствительные нервные окончания; 5-нервное волокно
Центральные хемочувствительные клетки реагируют на отклонения РСО2 и [H+] во внеклеточной жидкости внутримозгового интерстициального пространства, регулируют глубину вдоха. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы.
Одной из причин высокой скорости вентиляторного ответа на гиперкапнию является легкость диффузии СО2 через барьерную систему кровь-головной мозг. Более того, повышенное РСО2 вызывает расширение сосудов, особенно церебральных, способствуя тем самым усилению диффузии СО2 через гемато-энцефалический барьер.
Что отвечает за дыхание в головном мозге
8.1. Этиология и патогенез брадипноэ, гиперпноэ, тихипноэ, апноэ
Как известно, внешнее дыхание представляет собой процесс газообмена между внешней средой и альвеолярным воздухом, а также между альвеолами и притекающей к легким кровью (альвеолярное дыхание). Различают следующие виды нарушения внешнего дыхания:
Брадипноэ – редкое дыхание, которое возникает при поражении и угнетении дыхательного центра на фоне гипоксии, отека, ишемии и воздействия наркотических веществ, а также при перерезке n. vagus, несостоятельности хеморецепторного аппарата. При сужении крупных воздухоносных путей отмечается редкое и глубокое дыхание – стенотическое. Повышение артериального давления вызывает рефлекторное снижение частоты дыхательных движений (рефлекс с барорецепторов дуги аорты).
Гиперпноэ – глубокое и частое дыхание, отмечается при мышечной работе, гиперкапнии и высокой концентрации Н+, эмоциональном напряжении, тиреотоксикозе, анемии, ацидозе, снижении содержания кислорода во вдыхаемом воздухе (рефлекс с центральных и периферических хеморецепторов, проприорецепторов дыхательных мышц, с БАР воздухоносных путей и т.д.).
Тахипноэ – частое поверхностное дыхание. Возникает вследствие выраженной стимуляции дыхательного центра при гиперкапнии, гипоксемии и повышении концентрации ионов Н+(рефлекс с хемо-рецепторов, БАР и МАР воздухоносных путей и паренхимы легких). Этот вид дыхания наблюдается при лихорадке, пневмониях, застое в легких, ателектазе и т.п. Тахипноэ способствует развитию альвеолярной гиповентиляции в результате преимущественной вентиляции мертвого пространства.
Апноэ – отсутствие дыхательных движений наблюдается при гипокапнии, снижении возбудимости дыхательного центра вследствие поражения головного мозга, экзо- и эндоинтоксикаций, действия наркотических веществ.
Апнейстическое дыхание характеризуется судорожным усиленным удлиненным вдохом, который редко прерывается выдохом. Такой вид дыхания можно воспроизвести в эксперименте после перерезки у животного обоих блуждающих нервов и ствола на границе между верхней и средней третью моста.
Гаспинг-дыхание характеризуется редкими, глубокими, убывающими по силе “вздохами”, возникает в терминальной фазе асфиксии. При данном виде дыхания работают только клетки каудальной части продолговатого мозга.
8.2. Периодическое дыхание (Чейна-Стокса, Биота) и терминальное дыхание (Куссмауля)
Дыхание Чейна-Стокса характеризуется чередованием групп дыхательных движений с нарастающей амплитудой и периодов апноэ (остановка дыхания). Следует отметить, что дыхание Чейна-Стокса может встречаться не только в условиях патологии, но и в норме: у здоровых людей во сне, у недоношенных детей с незрелой системой регуляции дыхания, в старческом возрасте (когда имеется повышение порога возбудимости дыхательного центра).
Дыхание Биота характеризуется чередованием периодов апноэ с группами дыхательных движений равной амплитуды (рис.12).
В основе развития периодического дыхания Чейна-Стокса и Биота лежит снижение возбудимости дыхательного центра под влиянием экзогенных и эндогенных патогенных факторов, возникающее, в частности, при острой и хронической гипоксии, действии механической травмы мозга, развитии отека мозга, при аутоинтоксикациях (при сахарном диабете, почечной недостаточности, печеночной недостаточности), а также, на фоне развития коматозных состояний, воспалительного процесса, опухолевого поражения продолговатого мозга, при нарушениях мозгового кровообращения.
При дыхании Чейна-Стокса и Биота адекватные в условиях нормы гуморальные и нервные раздражители не оказывают стимулирующего влияния на бульбарный дыхательный центр Активация нейронов бульбарного дыхательного центра возникает лишь на фоне воздействия сверхпороговой афферентации с центральных и периферических хеморецепторов при возрастании РСО2, [H+] и снижении РаО2 по сравнению с показателями нормы.
В случаях развития дыхания Чейна-Стокса, несмотря на снижение возбудимости бульбарного дыхательного центра, возникает повышение активности бульбарных инспираторных нейронов на фоне дополнительной афферентации, возникающей в момент первых дыхательных движений с рецепторов воздухоносных путей, проприорецепторов дыхательных мышц и сухожильных рецепторов. Последнее приводит к углублению дыхания. Увеличение глубины дыхательных движений сопровождается нормализацией газового состава крови, когда дыхательный центр вновь оказывается нечувствительным к нормальным по силе гуморальным воздействиям. При этом вновь возникает апноэ, сопровождающееся развитием гипоксии, повышением концентрации РСО2 и [H+] сверх нормальных пороговых величин, вновь обеспечивающих гуморальную стимуляцию бульбарных инспираторных нейронов.
В случае развития дыхания Биота также имеет место снижение чувствительности нейронов бульбарного дыхательного центра к адекватным стимулирующим влияниям с хеморецепторов, их активация возникает лишь при воздействии сверхпороговых гуморальных раздражителей. Однако в случае развития дыхания Биота, в отличие от дыхания Чейна-Стокса, инспираторные нейроны дыхательного центра не реагируют повышением активности на дополнительную стимулирующую афферентацию, возникающую при первых дыхательных движениях с рецепторов воздухоносных путей, паренхимы легких, проприорецепторов дыхательных мышц и сухожильных рецепторов. В связи с этим амплитуда дыхательных движений не изменяется.
8.3. Одышка: виды и механизмы развития
1. Усиление стимулирующей афферентации в бульбарный дыхательный центр (с механорецепторов воздухоносных путей, с МАР растяжения альвеол, с рецепторов спадения, юкстакапиллярных рецепторов, проприорецепторов, а также с висцеральных рецепторов и рецепторов болевой, термической чувствительности).
2. Усиление рефлекторных влияний с барорецепторов аорты и каротидных синусов. Эти рецепторы включаются в патогенез одышки при кровопотере, шоке, коллапсе. При артериальном давлении ниже 70 мм рт. ст. (9,1 кПа и меньше) уменьшается поток импульсов, оказывающих тормозное влияние на центр вдоха.
3. При снижении в крови напряжения О2, увеличении СО2 или возрастании концентрации ионов [H+] возникают усиление афферентации с центральных и периферических хеморецепторов в бульбарный дыхательный центр, активация инспирации (сердечная недостаточность, дыхательная недостаточность, почечная недостаточность, анемии, нарушения КОС различного генеза).
4. Ощущение недостаточности дыхания может возникнуть при чрезмерном растяжении межреберных мышц (например, при тяжелой физической работе, уменьшении эластичности легких, сужении верхних дыхательных путей), когда возбуждаются проприорецепторы мышц, импульсация с которых поступает в высшие отделы головного мозга.
5. Стимуляция дыхательного центра и ощущение недостаточности воздуха имеет место при локальных нарушениях мозгового кровообращения (спазм, тромбоз сосудов мозга, эмболия, развитие деструктивных изменений инфекционного, аллергического и другого происхождения).
В случаях развития рестриктивных нарушений вентиляции легких (например, при бронхиальной астме, отеке легких, пневмофиброзе легочной ткани и т.д.), когда снижается эластичность легочной ткани и значительно затрудняется растяжение альвеол под влиянием физиологического объема вдыхаемого воздуха, с МАР растяжения альвеол длительное время поступает низкочастотная афферентация в бульбарный дыхательный центр. Последняя приводит к длительной стимуляции бульбарных инспираторных нейронов и удлинению акта вдоха, т.е. развитию инспираторной одышки.
При развитии обструктивной формы дыхательной недостаточности, в основе которой лежит нарушение проходимости воздухоносных путей, особенно в случаях развития бронхоспазма при атопической и инфекционной бронхиальной астме, возникает резкое затруднение удаления воздуха из альвеол в окружающую среду. При этом альвеолы длительное время находятся в перерастянутом состоянии. С МАР растяжения альвеол длительное время поступает высокочастотная импульсация в бульбарный дыхательный центр, пессимальная для инспираторных нейронов, что препятствует развитию акта вдоха и, соответственно, обеспечивает пролонгирование фазы экспирации, т.е. развитие экспираторной одышки.
Следует отметить, что выделение инспираторной и экспираторной одышки в определенной степени относительно, поскольку нередко одышка, начинаясь как инспираторная или экспираторная, достаточно быстро приобретает смешанный инспираторно-экспираторный характер.