Ультраструктура новой коры мозга горизонтальная и вертикальная организация
Толщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть.
Микроскопически кора имеет полосчатый, или пластинчатый, вид, отражающий распределение ее клеток и нервных волокон, а также веерообразную организацию клеточных элементов. Общая цитоархитектоника (схема строения, основанная на типе клеток; схему строения в зависимости от распределения миелинизированных волокон обозначают термином миелоархитектоника) во многом варьирует от зоны к зоне, что позволяет составить «карту» коры с десятками гистологически различных зон. Больших успехов удалось достичь в соотнесении этих зон со специфическими функциями. Несмотря на то, что это имеет теоретическую ценность, подобное разделение довольно упрощено, так как часто принимают во внимание только ключевые функции более обширных функциональных систем, взаимодействующих с другими отделами мозга.
а) Пластинчатое строение. Пластинчатое распределение нейронов характерно для всех отделов коры. Филогенетически «старые отделы», включая древнюю кору (обонятельную кору) и старую кору (гиппокампальная формация и зубчатое ядро; участвуют в процессах памяти) образованы тремя клеточными пластинками. Однако в новой коре (неокортексе или изокортексе, название которой указывает на единый нейрогенез коры, закончившийся образованием шестислойного строения), составляющей оставшиеся 90% (подавляющую часть коры больших полушарий), происходит переход к шестислойному строению.
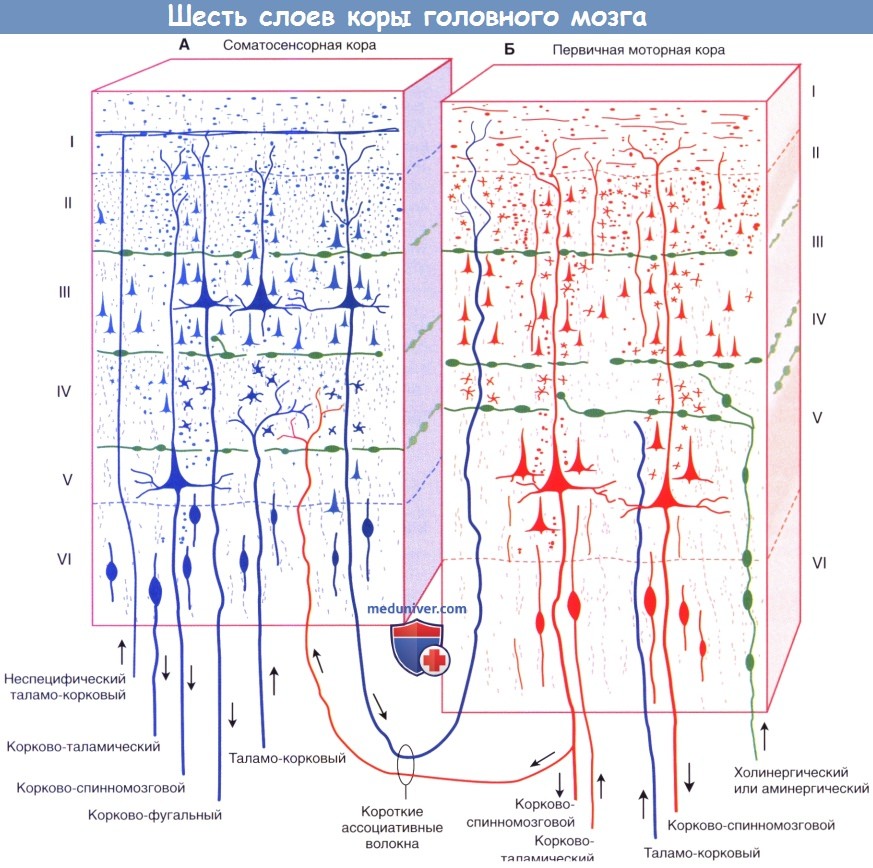 Шестислойное строение изокортекса коры головного мозга.
Шестислойное строение изокортекса коры головного мозга.
(А) Соматосенсорная кора. (Б) Первичная моторная кора, кортикальные пластинки обозначены цифрами от I до VI.
Клеточные пластинки новой коры (неокортекса):
I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса.
II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками.
III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток.
IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора.
V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу.
VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу.
б) Колончатое строение. Помимо хорошо различимой пластинчатой организации коры больших полушарий, можно наблюдать также лучистую, или «колончатую», организацию клеточных структур. Колончатое строение новой коры стало основой исследования функционирования групп нейронов в соматосенсорной коре животных. Подобные лучевидно расположенные группы клеток, вероятно, представляют собой отдельные зоны с одинаковыми физиологическими свойствами и образуют структурную основу для создания более сложных функций. Группы колонн могут образовывать модули, участвующие в обработке различных сенсорных модальностей и функций.
Установлено, что колонны в различных отделах коры имеют разное строение, обусловленное вариабельностью многочисленных свойств, включая строение и число клеток, онтогенез, синаптические связи и молекулярные маркеры. Все это влияет на различные функциональные характеристики и реакции в ответ на возбуждение. В качестве организующего принципа подобная концепция колончатого строения представляет определенную пользу, однако удобнее рассматривать кору как совокупность горизонтального (пластинчатого) и вертикального (лучистого) типов строения. Несмотря на то, что колонны не представляют собой отдельные подобные образования с четкими границами, такая концепция более точно отражает строение коры, а также наблюдаемые экспериментально функциональность, «экономию пространства» и пластичность коры.
Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи.
Лежащая в основе организации коры «схема» приводит к тому, что каждая из колонн становится модально (функционально) специфичной после «обработки» информации отдельными элементами. Однако конечная реакция проекционных нейронов колонн может значительно варьировать в зависимости от степени их возбуждения и информации, подходящей к каждому нейрону. Например, одна колонна может реагировать на движение в определенном суставе, но не на раздражение кожи над ним; однако при изменении условий их реакция также может изменяться.
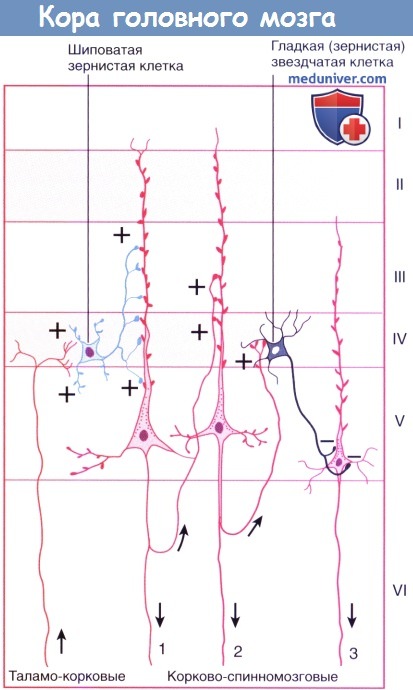 Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Пирамидная клетка 1 возбуждается шиповатой зернистой клеткой; она возбуждает клетку 2 внутри своей колонны клеток; клетка 3 в соседней колонне тормозится гладкой зернистой клеткой.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический.
Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.)
• Пирамидные клетки имеют пирамидоподобную форму с вершиной, направленной к поверхности. Размеры клеток в высоту составляют 20-30 нм в слоях II и III и более чем в два раза больше в слое V. Самые крупные — гигантские пирамидные клетки Беца — расположены в моторной коре. Единственный апикальный дендрит каждой пирамидной клетки достигает слоя I, заканчиваясь на пучке дендритов. Несколько базальных дендритных ветвей отходят от базальных «углов» клетки и веерообразно расходятся к соответствующим слоям. Апикальные и базальные дендритные ветви свободно разветвляются и усыпаны дендритными шипиками. Большинство пирамидных клеток расположено в слоях II-III и V-VI. Отходящие от основания клеток аксоны отдают несколько возвратных ветвей до вхождения в подлежащее белое вещество, их функция — возбуждение соседних пирамидных клеток.
• Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением.
Нейромедиатором различных по структуре и классификации непирамидных тормозных вставочных нейронов служит ГАМК. [Сложная классификация нейронов новой коры постоянно меняется. Гладкие звездчатые (зернистые) клетки можно обнаружить во всех слоях коры; их дендриты веерообразно расходятся во всех направлениях, а их аксоны образуют локальные древовидные разветвления, поэтому их иногда называют нейронами локальных сплетений. Несмотря на их уникальные морфологические характеристики, нейроглиеформные, клетки-канделябры и корзинчатые клетки считают специализированными типами зернистых клеток. Наш совет: если Вы встречаете термины «зернистая» или «гладкая звездчатая» клетка, для облегчения чтения и понимания их следует рассматривать как вставочные нейроны.]
Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR).
• Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях.
Корзинчатые клетки лежат преимущественно в слоях II и V, а свое название получили благодаря тому, что их аксоны формируют околоклеточные «корзинки» вокруг тел пирамидных клеток, дистальных отделов их дендритов и аксонов других корзинчатых клеток.
• Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств.
• 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
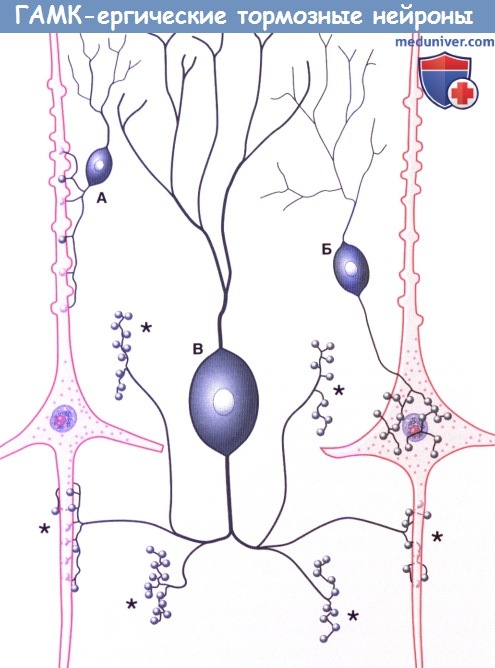 Три морфологических типа ГАМК-ергических тормозных нейронов:
Три морфологических типа ГАМК-ергических тормозных нейронов:
А. Аксодендритическая клетка, образующая синапс с апикальным отделом пирамидного нейрона.
Б. Корзинчатая клетка, образующая аксоаксональные синапсы с пирамидными клетками.
В. Клетка-канделябр, формирующая аксоаксональные синапсы (*) с начальными сегментами аксонов двух пирамидных клеток, показанных здесь, и с начальными сегментами четырех других клеток, не показанных на рисунке.
(На основе DeFelipe, 1999, с разрешения автора.)
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах.
1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария.
2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария.
3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках.
4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы:
• базальное ядро Мейнерта (базальные ядра мозга), ацетилхолин;
• серобугорно-сосцевидное ядро (задний отдел гипоталамуса), гистамин;
• черная субстанция, компактный слой (вентральный отдел покрышки среднего мозга), дофамин;
• ядра шва (средний мозг и ростральный отдел моста), серотонин;
• голубое пятно (ростральный отдел моста), норадреналин. Эти пять групп нейронов имеют особое значение в психиатрии и подробно рассмотрены в отдельной статье на сайте.
д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга.
• Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III.
• Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III.
• Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса).
Редактор: Искандер Милевски. Дата публикации: 22.11.2018
Ультраструктура новой коры мозга горизонтальная и вертикальная организация
В представленной работе сотрудники ФГБНУ «Научного центра неврологии» обобщили результаты последних исследований по вопросу структурных изменений, развивающихся в головном мозге при детском церебральном параличе. Анализ источников литературы позволил установить, что сроки воздействия этиологических факторов на развивающийся мозг плода определяют особенности поражения головного мозга и локализацию повреждения, а также характер ответа нервной системы.
Среди заболеваний нервной системы, выявляемых у детей, наиболее распространенное – детский церебральный паралич (ДЦП), который характеризуется клинически неблагоприятным течением и ранней инвалидизацией пациентов. Частота этого заболевания составляет 2–2,5 случая на 1000 новорожденных.
Согласно современным представлениям ДЦП – это группа перманентных нарушений, ограничивающих развитие двигательных навыков и рефлексов, формирующих позу ребенка, которые связаны с непрогрессирующим повреждением развивающегося головного мозга плода или новорожденного, часто сопровождаются расстройствами чувствительности и восприятия, а также нарушениями познавательной деятельности и поведенческих реакций. Повреждение головного мозга при ДЦП происходит задолго до рождения ребенка, а регресса симптоматики заболевания не наблюдается на протяжении всей жизни больного. Практикующие врачи (педиатры, неврологи, ортопеды и др.) сталкиваются в большинстве случаев с его остаточными проявлениями.
В этиологии заболевания ведущую роль отводят гипоксии, пери- и интравентрикулярным кровоизлияниям, перивентрикулярной лейкомаляции, внутриутробной инфекции и родовой травме.
Наряду с этим выделяют значительное число факторов риска, способствующих повреждению незрелого мозга плода: во внутриутробном периоде (неинфекционные заболевания у беременной женщины, прием антибиотиков, патология плаценты), при родах (акушерские пособия и кесарево сечение) и в раннем неонатальном периоде жизни ребенка (низкая оценка по шкале Апгар, свидетельствующая об асфиксии у новорожденного, и др.). Полиэтиологичность заболевания предполагает отсутствие единого подхода к объяснению механизмов его формирования или разнообразие самих механизмов, что косвенно подтверждается полиморфизмом клинических проявлений ДЦП: различают спастические, атактическую, дискинетическую и смешанную формы заболевания. Вместе с тем универсальность неспецифических механизмов, лежащих в основе патогенеза заболеваний нервной системы, обусловленная стандартным ответом нервной ткани при ее повреждении, позволяет обнаружить сходные структурные изменения в головном мозге детей с разными клиническими формами церебральных параличей.
Ультразвуковым методом у больных детским церебральным параличом в раннем возрасте в головном мозге выявляют порэнцефалические кисты и увеличенные размеры желудочков. Показано, что эти изменения связаны с кровоизлиянием в полость желудочков мозга. Наряду с этим ультразвуковое исследование головного мозга при детском церебральном параличе позволяет выявить снижение плотности перивентрикулярного белого вещества, что может быть обусловлено не только воздействием гипоксии, но и кровоизлиянием в эту область.
Методами МРТ установлено, что повреждение перивентрикулярной области возможно как на ранних этапах внутриутробного развития плода, так и в более поздние сроки. В этом случае так называемое окно уязвимости формируется в III триместре беременности, между 24-й и 34-й неделями внутриутробного развития. Предполагают, что в этом периоде вследствие повреждения свободными радикалами олигодендроглиоцитов, которые локализуются в перивентрикулярной области, нарушается формирование их миелиновых мембран.
На основе МРТ было также установлено, что у больных детским церебральным параличом повреждается не только перивентрикулярная область, но и нейронные структуры головного мозга. В зависимости от сроков повреждения при поражении серого вещества головного мозга у больных детей выявляют грубые пороки развития или умеренные дефекты.
Пороки развития головного мозга связывают с задержкой пролиферации нейронов, нарушением их миграции, нарушением образования архитектоники клеточных колонок и организации неокортекса. Миграция нейробластов в корковую пластинку (кору больших полушарий) начинается с ранних стадий ее развития. Кора больших полушарий головного мозга человека формируется по типу вертикальных, или радиальных колонок. Длинноаксонные нейроны – клетки пирамидного и веретеновидного типа, составляющие основу этих колонок (на ранних стадиях это нейробласты, мигрировавшие в корковую пластинку), начинают быстро развиваться задолго до дифференцировки коры на слои и миграции в нее короткоаксонных нейронов. Подобный тип развития корковой пластинки позволяет предположить, что вертикальные структуры коры больших полушарий мозга человека формируются раньше гори- зонтальных – слоев коры. Опережающая дифференцировка вертикальных структур коры может быть обоснована концепцией, согласно которой вертикальная организация коры отличается простотой строения – работает по типу классифицирующей машины и неспособна обучаться, что можно сопоставить с такими данными: к рождению ребенка вертикальные структуры коры (совокупность длинноаксонных нейронов) поля 4 характеризуются высокой степенью морфохимической дифференцировки, но двигательные условные рефлексы у ребенка еще не вырабатываются. Эти рефлексы начинают формироваться на основе ориентировочных реакций не раньше чем через 1,5 мес после рождения, а произвольные движения – с 3–4-го месяцев жизни. Следовательно, вертикальная колоночная организация сама не способна обеспечить работу коры на должном аналитико-синтетическом уровне. Для этого требуется созревание другой части единой системы – горизонтальной, т.е. слоев коры.
У больных ДЦП чаще выявляют такие пороки развития головного мозга, как голопрозэнцефалия, лиссэнцефалия, шизэнцефалия, пахигирия и полимикрогирия.
Подобного рода изменения чаще имеют двустороннюю и симметричную локализацию в правом и левом полушариях головного мозга и сопровождаются нарушением дифференцировки корковой пластинки на отдельные слои (при типичной микрогирии в поперечнике коры различают 4 слоя). Аналогичные пороки развития головного мозга регистрируют у больных, которые перенесли врожденную цитомегаловирусную инфекцию и впоследствии имели клинические проявления детского церебрального паралича.
Умеренные дефекты в структурах головного мозга чаще описывают как атрофические изменения различных отделов корковой пластинки и очаги глиоза, которые формируются в III триместре внутриутробного развития. К этому же периоду относят поражения подкорковых ядер. Поражение подкорковых ядер, в частности чечевицеобразного ядра, и минимальные изменения в корковой пластинке обнаруживают при неспастических формах ДЦП, тогда как генерализованные корково-подкорковые поражения, сопровождающиеся морфологическими изменениями в височной области коры, гиппокампе, подлежащем белом веществе и других образованиях головного мозга, наблюдается при спастических формах этого заболевания, осложненных эпилепсией. Поражения мозжечка у больных детским церебральным параличом регистрируют в виде инфарктов, симметричных и асимметричных атрофических изменений, нередко сочетающихся с увеличением IV желудочка.
Кроме того, показано, что в неонатальном периоде наиболее часто структурные изменения в головном мозге определяют в связи с развитием перинатальных артериальных ишемических инсультов, которые в 39% случаев приводят к формированию ДЦП. Методами МРТ регистрируют инфаркты в области внутренней сонной артерии, передней и средней мозговых артерий. Наряду с этим у больных выявляют глубокие инфаркты серого вещества головного мозга.
Другим типом инфаркта головного мозга, который наблюдается при ДЦП, является неонатальный венозный перивентрикулярный инфаркт. По распространенности он не только не уступает артериальным ишемическим инсультам, но и превышает их (в соотношении 62 : 44). С этим типом инфаркта связывают формирование гемиплегической формы заболевания.
С помощью МРТ можно получить представление о локализации очага повреждения и наличии атрофии нейронных структур в образованиях головного мозга, но этот метод не позволяет определить объем той или иной структуры у каждого пациента и сопоставить его с аналогичными показателями в контрольной группе. Подобную возможность предоставляет воксел-ориентированная морфометрия – метод компьютерного моделирования, основанный на трехступенчатой процедуре выравнивания, т.е. наложения каждого МРТ-среза на соответствующее сканированное изображение. При помощи этого метода у пациентов с ДЦП по сравнению с контрольной группой выявили уменьшение объема серого вещества в передней и задней центральных извилинах, парацентральной дольке и зрительной области коры головного мозга. Этим же методом у детей с церебральными параличами регистрировали уменьшение объема серого вещества в островке мозга, таламусе, подкорковых ядрах (чечевицеобразном ядре и ограде) и левой ножке мозга.
Вместе с тем для прогнозирования возможных последствий повреждения головного мозга в перинатальном периоде данных, полученных методами функциональной диагностики и МРТ, явно недостаточно, так как они не дают полной картины повреждений.
Наиболее объективно процесс повреждения головного мозга отражают патологоанатомическое и патогистологическое исследование аутопсийного материала. На аутопсийном материале у детей с ДЦП было выявлено поражение большинства образований головного мозга. На макроскопическом уровне это проявлялось уменьшением размеров полушарий большого мозга и полушарий мозжечка, атрофией лобных и затылочных долей, полимикрогирией, истончением мозолистого тела, уменьшением размеров базальных ядер и таламуса, размягчением белого вещества. Морфологические изменения, определяемые при патологоанатомическом исследовании у больных ДЦП, существенно отличались от изменений, выявляемых у детей, перенесших черепно-мозговую травму в возрасте 2–3 мес и умерших впоследствии в промежутке от 2,5 года до 17 лет.
Гистологическое исследование, проведенное у больных детским церебральным параличом, позволило обнаружить в белом веществе перивентрикулярной области цитотоксический отек, вакуолизацию глиальных элементов, пикноз их ядер и набухание аксонов. Кроме того, отмечали реакцию астроглии и макрофагов. Астроглию локализовали при помощи иммуногистохимических реакций на выявление виментина и кислого глиофибриллярного белка GFAP. Астроцитарная реакция выражалась в формировании глиального рубца, отграничивающего очаги лейкодистрофии, и в наличии некроза в белом веществе. Макрофаги определяли реакцией на микроглиальный маркер Iba1 и маркеры макрофагов CD45, CD68 и др. В результате обнаружили, что увеличение числа макрофагов сопровождалось накоплением в них липидов. Клетки микроглии, представляющие собой иммунокомпетентные макрофаги, мгновенно реагируют на мельчайшие изменения в нервной ткани при патологических состояниях, фагоцитируя поврежденные нейроны и сокращая количество аксонов.
В двигательной области коры больших полушарий (поля 4 и 6) головного мозга у больных ДЦП находили очаговое выпадение типичных пирамидных нейронов в III и V слоях и, напротив, сохранение клеточных элементов IV слоя, что нехарактерно для агранулярной коры. Кроме того, в двигательной области коры обнаруживали множественные нейронные гетеротопии. При изучении нейроглиальных взаимоотношений выявляли уменьшение плотности расположения нейронов в области ядер черепных нервов, начиная со второго года жизни. В зрительной области коры (поле 17) во всех слоях корковой пластинки у больных определяли перицеллюлярный отек. У больных детей первого года жизни II–IV слои не были четко дифференцированы и выглядели как сплошной клеточный конгломерат. В I и IV слоях корковой пластинки были выявлены гетеротопии нервных клеток. Плотность расположения нейронов в основных слоях поля 17 зрительной коры у детей с детским церебральным параличом существенно отставала от контроля, тогда как плотность расположения глиоцитов начиная с 3 мес жизни превышала контрольные величины.
Морфологические изменения у детей с ДЦП, обнаруженные в двигательной и зрительной областях коры головного мозга (недостаточная дифференцировка слоев верхнего этажа корковой пластинки, отставание от нормы показателя плотности расположения нейронов, значительный глиоз), коррелировали с изменениями, выявленными в коре теменной области мозга у детей с тяжелыми формами другого заболевания – врожденной гидроцефалии. Учитывая врожденный характер этих заболеваний, можно предположить, что незавершенность формирования клеточных слоев корковой пластинки в обоих случаях могла быть связана как с морфофункциональной незрелостью нервных клеток, так и с нарушением дифференцировки корковой пластинки на отдельные слои. Указанные изменения могли быть обусловлены воздействием на плод гипоксии или инфекции, т.е., они отображали неспецифические реакции нервной ткани в ответ на воздействие неблагоприятных факторов.
Недостаточная дифференцировка слоев верхнего этажа корковой пластинки у детей с ДЦП свидетельствует о повреждении у них головного мозга на этапе дифференцировки поперечника коры на отдельные слои, которое завершается на 25–27-й неделе внутриутробного развития, а изменения в местоположении у них нервных клеток (гетеротопия) предполагает нарушение миграции нейронов в плащевой отдел (миграция протекает с 14-й по 23-ю неделю внутриутробного развития).
Таким образом, обобщая изложенное, можно сделать вывод, что при ДЦП повреждение головного мозга может происходить на разных этапах внутриутробного развития плода.
В одних случаях оно наблюдается в период миграции нейробластов вдоль отростков радиальной нейроглии в плащевой отдел (нейронные гетеротопии), в других – на этапе развития и дифференцировки нейронов (отставание в размерах тел нейронов). Третий вариант – повреждение головного мозга на этапе дифференцировки поперечника корковой пластинки на отдельные слои (слабая дифференцировка нейронов верхнего этажа коры, сохранение IV слоя в агранулярной коре), четвертый вариант – повреждение на этапе формирования борозд и извилин (признаки корковой дисплазии – лиссенцефалия, шизэнцефалия и др.). В остальных случаях головной мозг у детей поражается в более поздние сроки, чаще в последние месяцы внутриутробного развития, когда ткани плода реагируют воспалительной реакцией в ответ на воздействие неблагоприятных факторов. Следовательно, этап внутриутробного развития, во время которого происходит неблагоприятное воздействие, определяет особенности поражения головного мозга и локализацию повреждения, а также характер ответа нервной системы.
Сальков В.Н., Худоерков Р.М.
Российский вестник перинатологии и педиатрии, 2019; 64:(3)