В головном мозге рептилий ведущую роль берет на себя
Пресмыкающиеся – это настоящие сухопутные животные, которые могут жить, размножаться и развиваться вдали от водоемов. Они относятся к высшим позвоночным. Их нервная система в связи с подвижным и сложным образом жизни развита лучше, чем у амфибий.
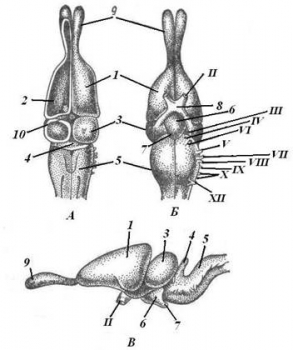
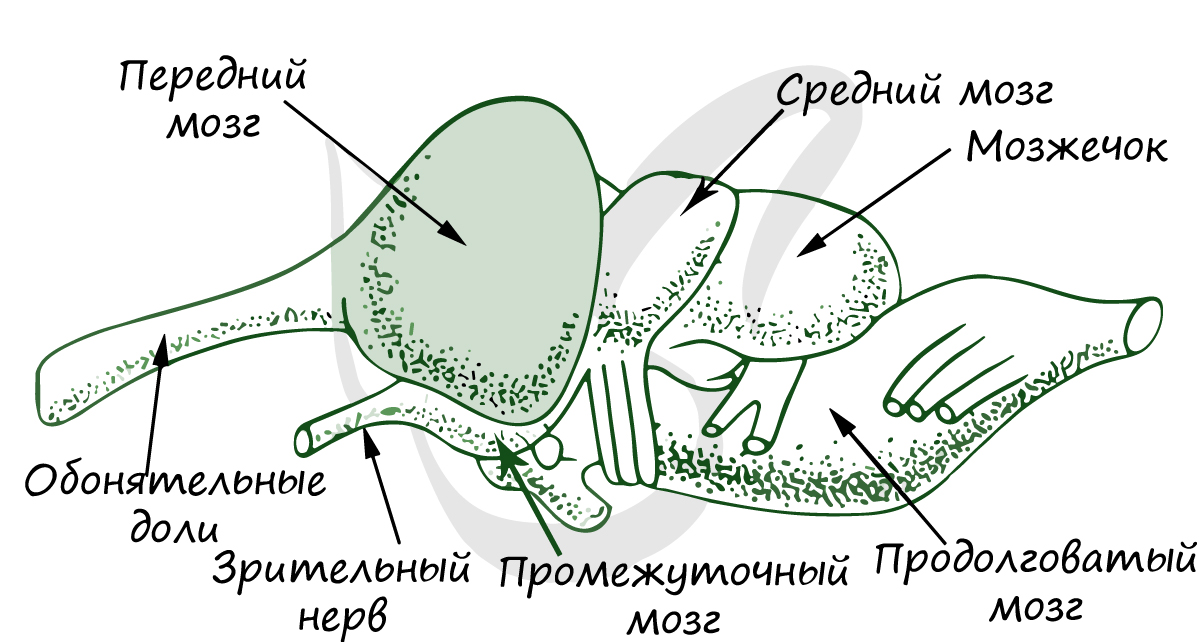
Передний мозг значительно больших размеров, чем у земноводных и имеет более сложное строение; у них возрастает способность к образованию условных рефлексов, быстрее устанавливаются новые связи с внешней средой и они лучше своих предков могут приспосабливаться к изменениям среды. Передний мозг состоит из двух полушарий, которые разрастаясь назад, прикрывают промежуточный мозг за исключением эпифиза и теменного органа. Увеличение переднего мозга происходит в основном за счет полосатых тел (скопления нейронов), располагающихся в области дна боковых желудочков. Они выполняют роль высшего интегративного центра, обеспечивая анализ поступающей в передний мозг информации и выработку ответных реакций. Таким образом, он перестает быть только обонятельным центром. Подобный тип мозга получил название зауропсидного. Что касается мозгового свода, то в нем происходят важные преобразования. В обоих полушариях крыши переднего мозга впервые в эволюции появляется по два островка серого вещества (зачатки коры) – один из них располагается на медиальной, а другой на латеральной стороне полушарий. Функционально значимым является только медиальный островок, который представляет собой высший обонятельный центр. В целом островки коры имеют примитивное строение и называются древней корой (archicortex). Большинство авторов считают островки коры однослойными, хотя у крокодилов можно выделить два и даже три слоя.
Связанные с передним мозгом обонятельные доли хорошо развиты. У одних видов они занимают сидячее положение, но чаще дифференцированы на луковицу и стебель.
Исследование переднего мозга рептилий имеет важное значение для эволюционной нейрогистологии, т.к. они являются ключевой точкой в эволюции позвоночных животных, начиная от которой развитие переднего мозга пошло по двум принципиально разным направлениям: по стриарному пути с преимущественным развитием подкорковых структур к птицам и по кортикальному пути с преимущественным развитием корковых структур к млекопитающим.
Промежуточный мозг на тонкой крыше имеет два пузыревидных образования, одно из которых располагается впереди и называется теменным, или парапинеальным, органом, а второе позади – это эпифиз (пинеальная железа). Парапинеальный орган выполняет светочувствительную функцию, и поэтому его еще называют теменным глазом. По сути парапинеальный орган и эпифиз составляют тандем, который является регулятором суточной активности животных. Однако теменной орган встречается не у всех рептилий. В таких случаях механизм регуляции суточной активности будет другой: информация о длине светового дня поступает не от парапинеального органа, а от зрительной системы.
Средний мозг представлен двухолмием и в основных чертах имеет ту же организацию, которая свойственна и амфибиям, однако для рептилий характерно более точное топографическое представительство в среднем мозге каждой из сенсорных систем. Кроме того, практически все моторные ядра мозжечка и продолговатого мозга взаимодействуют с нейронами крыши среднего мозга. Вместе с тем средний мозг утрачивает значение основного интегративного отдела центральной нервной системы. Эта функция переходит к переднему мозгу.
Часть зрительных и слуховых волокон по обходным путям, минуя средний мозг, направляется в передний. В среднем мозге при этом сохраняются центры обеспечения автоматических врожденных реакций организма, полученных еще на ранних этапах эволюции позвоночных. Новые центры переднего мозга принимают на себя функции текума и формируют новые двигательные пути.
Мозжечок в связи с освоением рептилиями ходьбы и бега развит лучше, чем у амфибий. Он состоит из центральной осевой части, называемой червем, а у некоторых намечаются и боковые лопасти. Для мозжечка характерны многочисленные связи с другими отделами нервной системы, имеющими отношение к локомоции. Относящийся сюда спиномозжечковый тракт, присутствующий и у рыб и у амфибий, распространяется и на дорзальный отдел червя. Имеют место мозжечковые связи с вестибулярным аппаратом, средним и ромбовидным мозгом. Вестибулярно-мозжечковые связи осуществляют контроль за положением тела в пространстве, а таламические регулируют мышечный тонус.
Продолговатый мозг образует резкий изгиб в вертикальной плоскости, характерный вообще для амниот.
От ствола мозга отходит 12 пар черепно-мозговых нервов.
Класс пресмыкающиеся
На настоящий момент класс насчитывает около 10 тысяч видов.


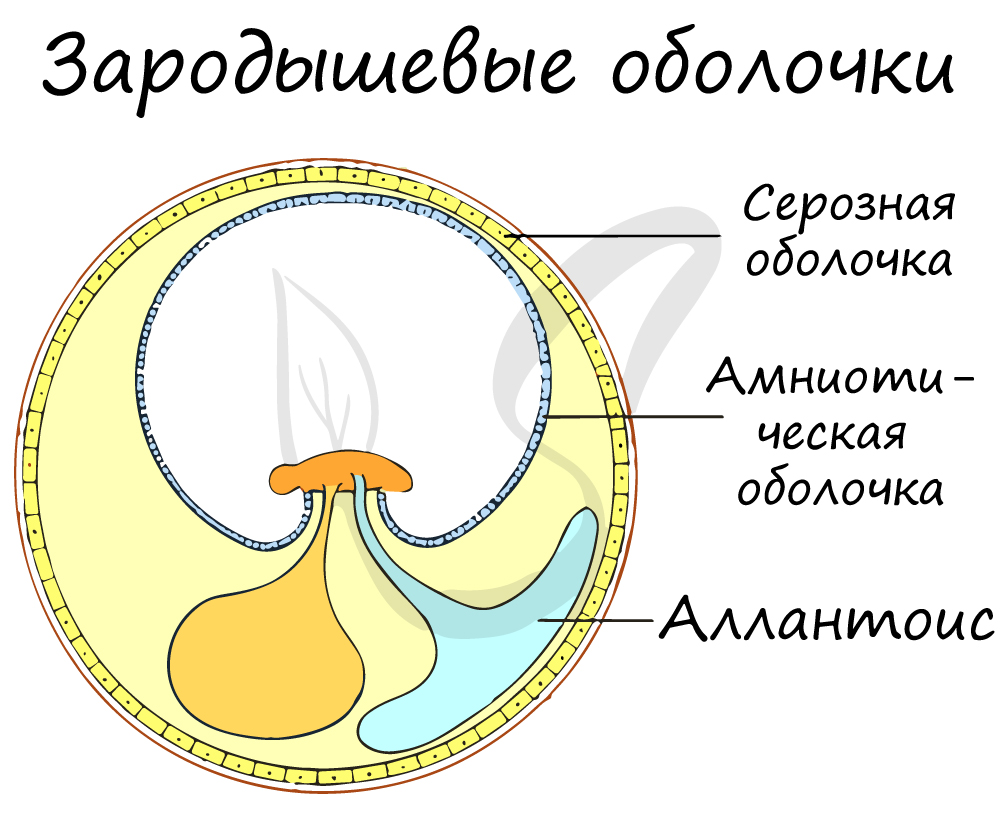
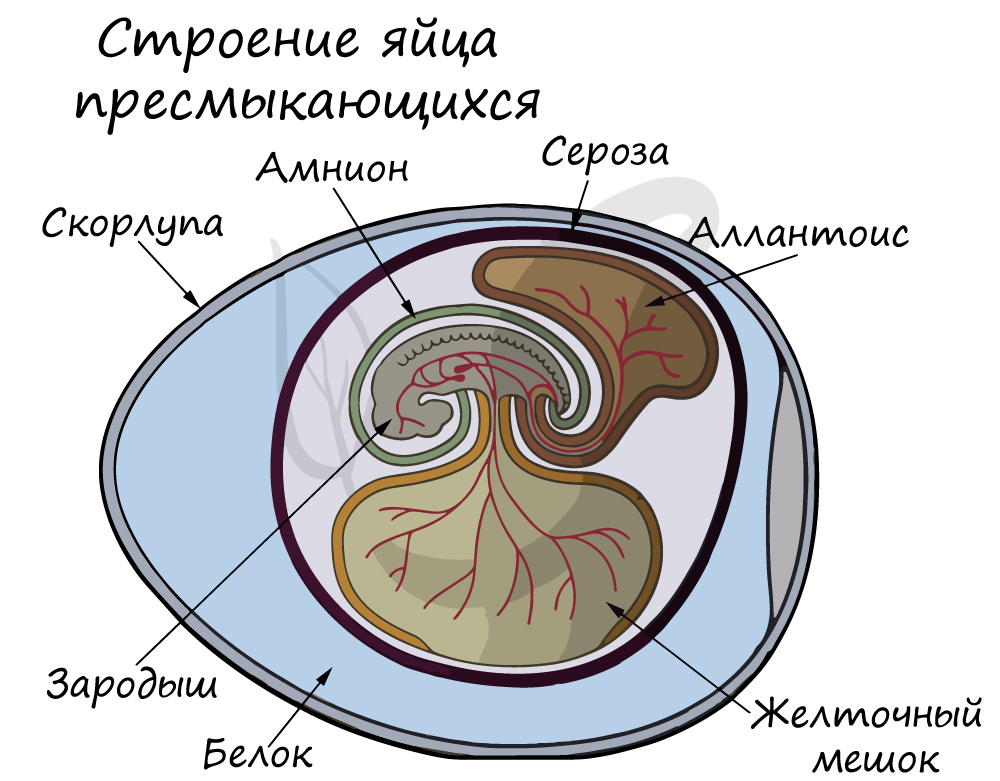
У рыб и земноводных оплодотворение было наружным, малоэффективным, с небольшой вероятностью встречи половых клеток. При внутреннем оплодотворении, которое появляется у рептилий, семенная жидкость самца со сперматозоидами вводится непосредственно в половые пути самки, что значительно увеличивает вероятность встречи мужских и женских гамет.

Кожа пресмыкающихся сухая, практически лишена желез (которых так много у земноводных). Верхние слои эпителия ороговевают, на поверхности кожи формируются роговые чешуйки. Эта защита необходима от пересыхания, помогает эффективнее удерживать воду в организме.

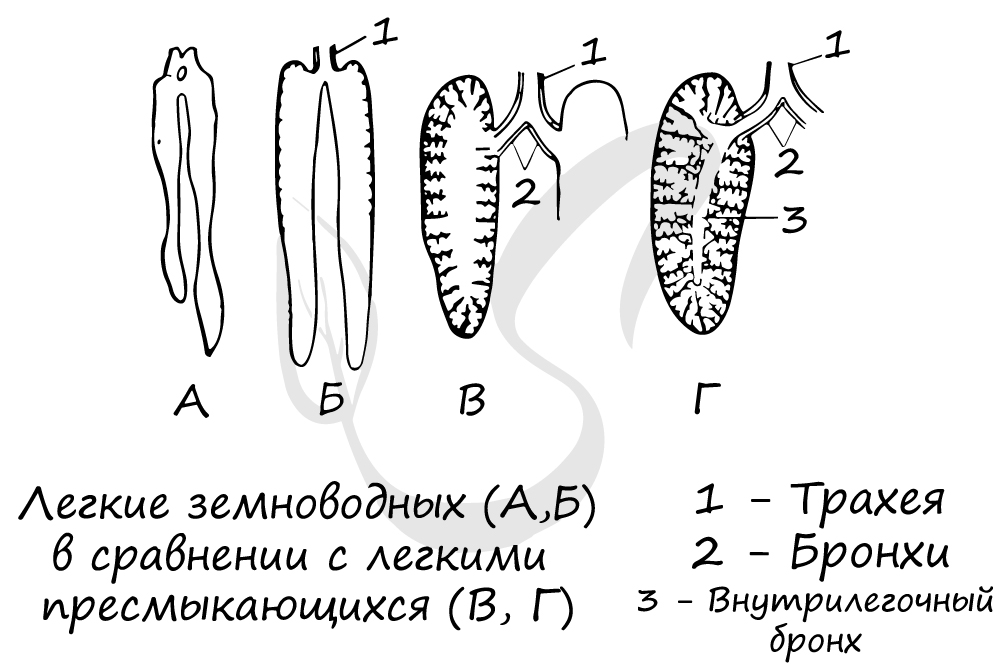
У пресмыкающихся происходит усложнение строения дыхательной системы: появляются ячеистые легкие, обеспечивающие более эффективный газообмен. Кожное дыхание (доминировавшее у амфибий) у пресмыкающихся сведено к минимуму или отсутствует вовсе.
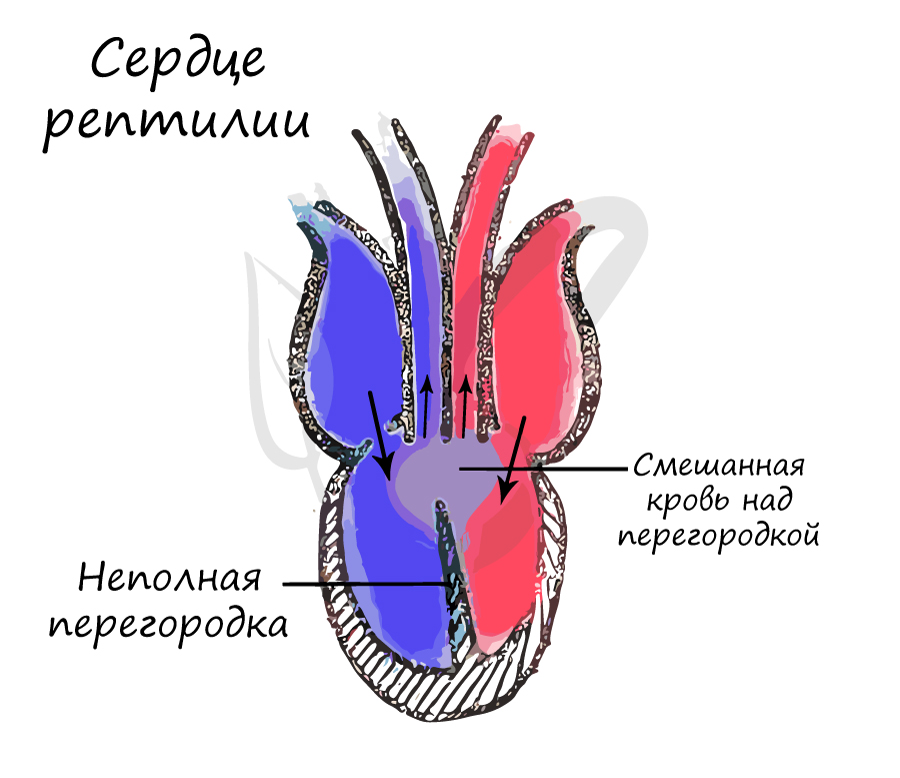
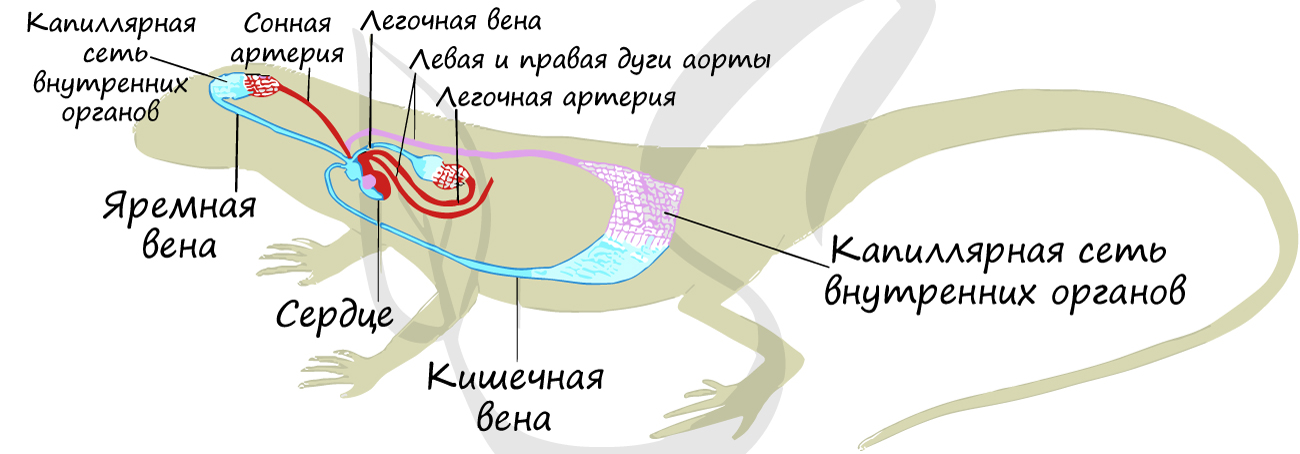
Тем не менее, над перегородкой кровь смешанная, так что пресмыкающиеся, как и земноводные, относятся к пойкилотермным (холоднокровным) животным.
У пресмыкающихся впервые возникают зачатки новой коры головного мозга, совершенствуется воспринимающая и интегрирующая функции головного мозга. Становится возможным более сложное поведение.
Отряды пресмыкающихся

Тело ящерицы покрыто сухой кожей, практически лишенной желез, с роговыми чешуями и щитками. Такое строение покровов тела предотвращает высыхание организма, защищает от потери воды. Испарение через кожу сохраняется, но в минимальном объеме.

Тело подразделяется на голову, туловище, хвост и две пары конечностей. Конечности расположены по бокам туловища, поэтому поднять голову высоко над землей пресмыкающиеся не могут. Конечности пятипалого типа, перепонки между пальцами отсутствуют.
В случае нападения хищника автотомия может спасти жизнь ящерицы, так как отброшенный хвост приковывает к себе внимание и хищник перестает преследовать ящерицу.

К ребрам крепятся впервые возникшие у пресмыкающихся межреберные мышцы, участвующие в дыхании.

Скелеты поясов конечностей служат опорой для самих конечностей. Передний (плечевой) пояс состоит из парных ключиц, лопаток, вороньих костей (коракоидов) и надгрудинника (надключицы), а также хрящей. Задний (тазовый) пояс конечностей образован тремя костями: седалищной, лобковой и подвздошной, также включает в себя хрящевые прослойки между ними.
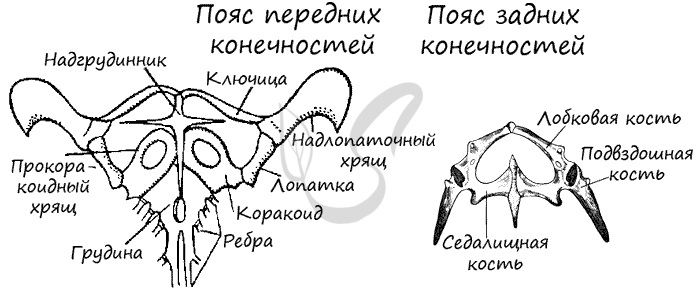
Скелеты передних и задних свободных конечностей аналогичны по строению конечностям амфибий. Однако стоит заметить, что голень и предплечье не состоят из одной единой кости: они подразделяются на лучевую и локтевую (составляют предплечье), малоберцовую и большеберцовую (составляют голень).

Строение пищеварительной системы рептилий более дифференцировано по сравнению с таковым у земноводных.
Ротовая полость переходит в короткую глотку, которая продолжается в пищевод. Пищевод расширяется и переходит в желудок, от которого начинается тонкий, а затем толстый отдел кишечника. В тонкую кишку открываются протоки печени, поджелудочной железы и желчного пузыря.
Граница между тонкой и толстой кишкой особенно хорошо выражена: на ее месте располагается зачаток слепой кишки. Оканчивается пищеварительная система клоакой.
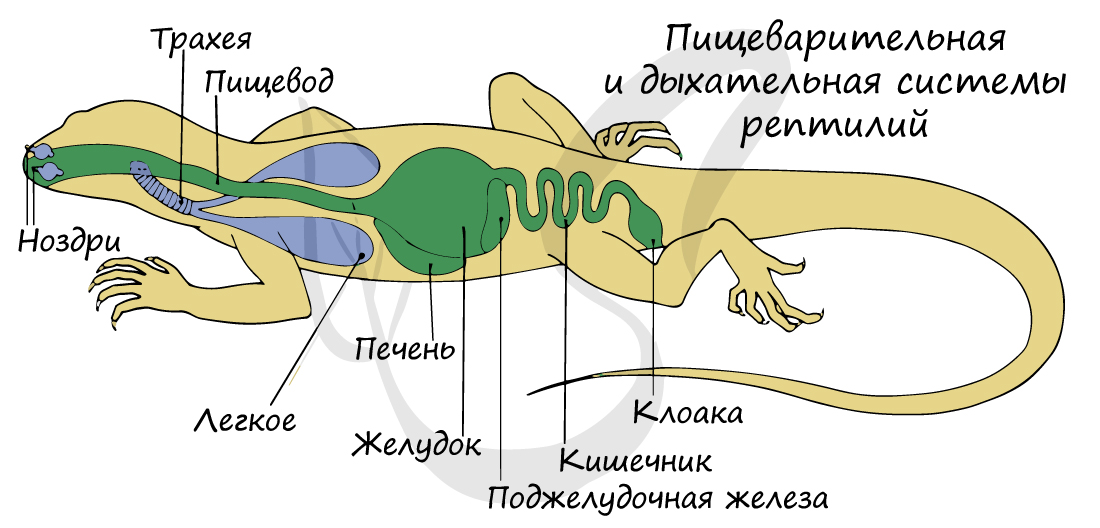
Кожа не участвует в дыхании, эту функцию берут на себя возникшие у рептилий ячеистые легкие. По сравнению с легочными мешками амфибий, ячеистые легкие имеют бо́льшую дыхательную поверхность.
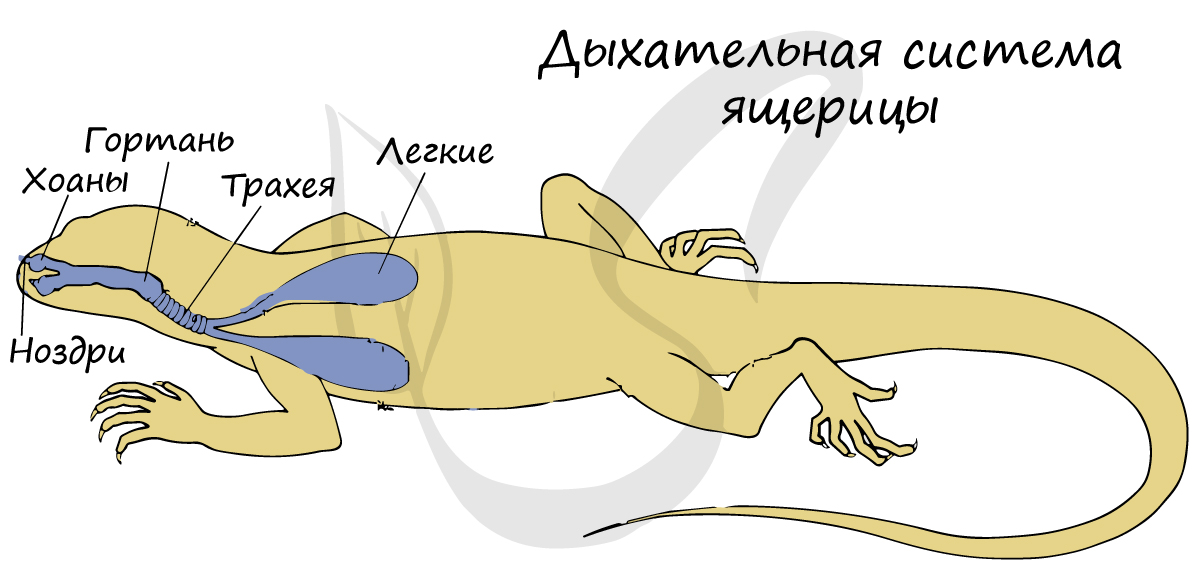
Поскольку рептилии холоднокровные животные, то интенсивность их обмена веществ напрямую зависит от температуры окружающей среды. Частота дыхательных движений также коррелирует с метаболизмом, поэтому чем выше температура окружающей среды, тем чаще дышит животное.
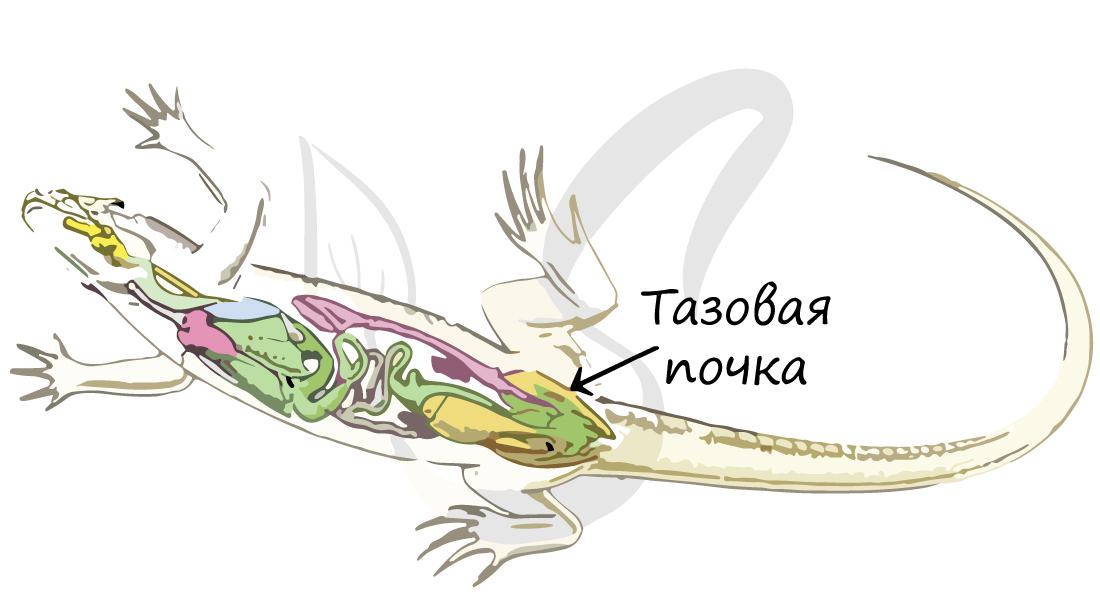
Сравнивая головной мозг земноводных и пресмыкающихся, становятся заметны несколько существенных отличий.
Мозжечок развит лучше, чем у земноводных. У рептилий появляются зачатки новой коры переднего мозга, он значительно увеличивается в объеме и перестает быть исключительно обонятельным центром, вбирая в себя новые функции.
Для рептилий характерно более сложное (чем для амфибий) поведение, помимо заложенных генетически безусловных рефлексов, они способны вырабатывать адаптивные, условные рефлексы.
Веки и мигательная перепонка служат для увлажнения поверхности глаза и предохраняют от высыхания.

Орган слуха также совершенствуется: появляется круглое окно во внутреннем ухе, улучшается передача звуковых колебаний, благодаря чему слух рептилий становится острее. Среднее ухо по-прежнему представлено стремечком и барабанной перепонкой.

Мужская половая система представлена семенниками и отходящими от них семяпроводами, впадающими в мочеточники. Мочеточники открываются в клоаку. У самцов имеются особые совокупительные органы, предназначенные для введения семенной жидкости в половые пути самки. Оплодотворение у рептилий внутреннее.
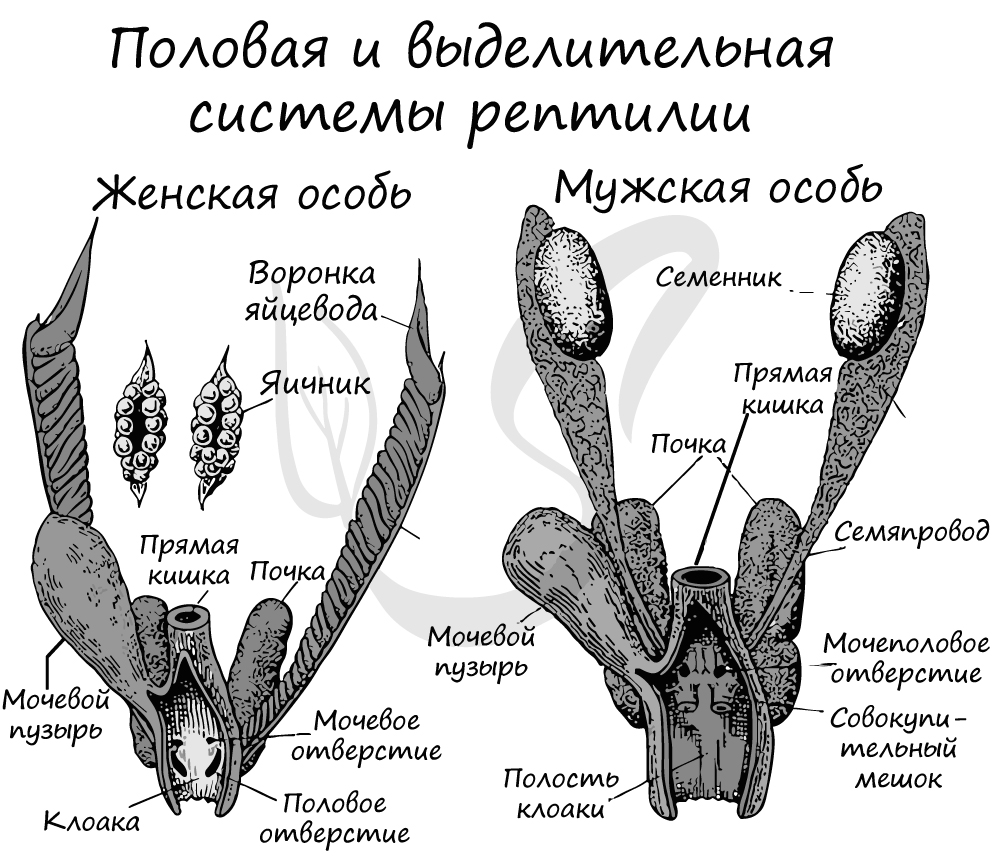
У самок половая система состоит из яичников, от которых отходят яйцеводы, впадающие в клоаку. Сперматозоиды оплодотворяют яйцеклетки в верхней части яйцевода.

Змеи также принадлежат к отряду чешуйчатые. Это безногие животные, адаптировавшиеся к перемещениям в густом лесном покрове, кронах деревьев. У змей взгляд немигающий: отсутствуют подвижные веки. Однако их отсутствие не означает, что поверхность глаз не нуждается в увлажнении: у них имеется мигательная перепонка, смачивающая поверхность глаза.

Змеи подразделяются на ядовитых и неядовитых. Ядовитыми являются гадюка обыкновенная, песчаная эфа, черная мамба, кобра, грюза, гремучие и морские змеи. К неядовитым относятся полозы, ужи, удавы.

Черепахи
Имеют уплощенное тело, покрытое панцирем из двух щитков: брюшного и спинного. Примечательно, что на задних конечностях имеются межпальцевые перепонки для плавания. Морские черепахи продвинулись еще дальше: у них задние конечности видоизменены в ласты, которые служат приспособлением к водному образу жизни.

Крокодилы
Однако, крокодилы все равно остаются холоднокровными животными: кровь смешивается в спинной аорте, образованной в результате слияния левой и правой дуги аорты.

Клювоголовые

Значение пресмыкающихся
Как и все живые организмы, пресмыкающиеся являются звеном в цепи питания (консументами). Регулируют численность насекомых, питаясь ими. Среди рептилий встречаются как хищники, так и растительноядные формы.
Рептилии имеют пищевое значение. В Африке и Азии употребляют в пищу около сотни различных блюд из мяса змей; яйца, жир и мясо морской черепахи также употребляются в пищу. Из кожи крокодилов изготавливают различные изделия.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Рептилии (Reptilia)
Автор
Редактор
Рептилии (Пресмыкающиеся) являются ключевым классом высших позвоночных животных, от которых эволюция пошла по пути формирования двух других классов высших позвоночных – птиц и млекопитающих. Рептилии – первый класс позвоночных, полностью вышедших на сушу (водные черепахи, морские змеи – вторично освоили водную среду), что привело к кардинальным изменениям во всех системах органов животных, включая нервную систему и органы чувств. С позиции эволюционной нейроморфологии рептилии представляют исключительный интерес, поскольку именно в этом классе позвоночных закладываются те особенности организации головного мозга, которые дали начало развитию высших центров мозга птиц и млекопитающих.
К этим особенностям относятся:
Содержание
Общий обзор строения нервной системы Рептилий
Спинной мозг
(Medulla spinalis)
У рептилий в спинном мозге происходит дальнейшая дифференциация серого и белого вещества. Особенно это касается распределения групп мотонейронов в передних рогах серого вещества. Мотонейроны спинного мозга рептилий организованы в две моторные группы: вентромедиальную и вентролатеральную, нейроны которых отличаются друг от друга по размеру и характеру ветвления дендритов. У рептилий впервые среди позвоночных дендриты большинства мотонейронов не проникают на противоположную сторону спинного мозга, и это позволяет осуществлять более тонкую регуляцию деятельности мотонейронов спинномозгового сегмента через систему интернейронов (рис. 2).
Мотонейроны первой группы иннервируют осевую мускулатуру, второй – мускулатуру конечностей. У рептилий мотонейроны, иннервирующие отдельные мышцы, уже расположены раздельно, тогда как у амфибий мотонейронные пулы разных мышечных групп перемешаны. Кроме того, у рептилий (особенно у змей) самая сложная среди позвоночных мышечная система, в связи с чем, у них встречается несколько вариантов моторной иннервации. Есть мышцы, которые иннервируются мотонейронами, расположенными в разных сегментах спинного мозга. Такая иннервация называется полинейронной, причем нейромышечные контакты от разных мотонейронов располагаются по всей длине мышечного волокна. Другие мышцы иннервируются мононейронально и локально. Очень часто в отдельных мышцах красные и белые волокна перемешаны друг с другом и иннервируются разными мотонейронами. Считается, что такая иннервация мышц у рептилий обеспечивает синхронизацию их работы и более тонкую регуляцию (например, при движении туловища у змей).
Все проводящие пути спинного мозга сосредоточены в белом веществе, разделяемом на три канатика: задний, боковой и передний. Основной объем канатиков занимают супраспинальные тракты, осуществляющие двустороннюю связь спинного и головного мозга (спинно-ретикулярные, спинно-тектальные, спинно-таламические, спинно-мозжечковые и др.). В отличие от низших позвоночных, у рептилий намечается четкая дифференциация проводящих пучков. Впервые выделяется зона проприоспинальных трактов. Нисходящие супраспинальные тракты (ретикуло-, вестибуло-, текто-, руброспинальные и др.) опосредуют влияние головного мозга на моторные центры спинного мозга и имеют сходное строение у всех позвоночных.

Рис. 1. Общий вид головного мозга некоторых представителей современных рептилий (по: Обухов, Андреева, 2017; Bangma, 1983).

Рис. 2. Нейронная структура вентромедиальной (а) и вентролатеральной (б) группы мотонейронов в спинном мозге ящерицы (по: Cruce, 1979).
1 и 2 – задние и передние, соответственно, рога серого вещества спинного мозга; 3 – медиальные проприоспинальные тракты; 4 – спинномозговой канал; VII–IX – пластины Рекседа

Рис. 3. Схема строения и эволюции мозжечка наземных позвоночных животных (по: Bangma, 1983).
а – амфибии, б – рептилии, в – птицы, г – млекопитающие, 1 – зачаток червя мозжечка (vermis), 2 – паравермальная зона, 3 – тело (полушария) мозжечка, fl – клочок
Продолговатый мозг
(Medulla oblongata)
Спинной мозг (Medulla spinalis) имеет сходное строение со всеми позвоночными. Основным принципом организации спинного мозга является его сегментарное строение и внутреннее разделение на серое и белое вещество. Он располагается в позвоночном канале от первого шейного позвонка до крестца, где переходит в краевую нить. По ходу спинного мозга имеется два утолщения: шейное, сегменты которого управляют крыльями и пояснично-крестцовое, связанное с иннервацией задних конечностей. Разделение серого вещества на передние (вентральные) и задние (дорсальные) рога у птиц выражено слабо. Боковые рога практически отсутствуют, в связи с чем, нейроны, связанные с вегетативной нервной системой, располагаются ближе к центральному району серого вещества (рис. 1).
Формирующиеся у рептилий и других высших позвоночных связи сенсорных ядер продолговатого мозга с моторными и вегетативными центрами головного мозга обеспечивают такие сложные рефлекторные реакции как жевание, глотание, слюноотделение, рвотный рефлекс. Восходящие связи сенсорных областей мозга связаны с формированием пищевого и питьевого поведения. Наибольшие изменения в продолговатом мозге рептилий связаны с октаво-латеральной системой. Переход к наземному образу жизни привел к редукции центров октаво-латеральной системы, связанных с боковой линией и электрорецепцией и последующим развитием вестибулярных и слуховых центров (VIII – слухо-статический нерв). Формируется сложный комплекс вестибулярных и слуховых ядер, связанных трактами со спинным и средним мозгом. Вентральные отделы продолговатого мозга занимает висцеромоторная зона, которая включает моторные ядра тройничного, лицевого, языкоглоточного и блуждающего нервов. Они обеспечивают иннервацию дыхательной и языко-глоточной мускулатуры. Соматомоторная зона связана с моторными центрами III, IV, VI (глазодвигательные ядра) и XII (подъязычный нерв) черепно-мозговых нервов. Интересно, что среди рептилий наиболее сложным строением моторного ядра XII нерва обладают хамелеоны, отличающиеся наиболее сложным мышечным аппаратом языка. Таким образом, у рептилий, при сохранении общего плана строения продолговатого мозга, наблюдается четкая эволюционная тенденция к усложнению и дифференциации как морфологической структуры, так и системы проводящих трактов мозга.
Задний мозг (Metencephalon) и мозжечок (Cerebellum)
У рептилий в мозжечке выделяют традиционные для всех позвоночных отделы: латеральные области и медиальную часть. Латеральные области, хорошо развитые у низших позвоночных и связанных с октаво-латеральной системой и получившие наименование ушки мозжечка (auriculae cerebelli) у рептилий практически редуцируются и превращаются в клочково-узелковую долю (lobus flocculo-nodularis). Медиальная часть составляет тело мозжечка (corpus cerebelli), разделяемый бороздами на доли и дольки (рис. 3). В глубине тела мозжечка у рептилий формируются два ядра, нейроны которых образуют систему эфферентных связей.
Гистологическая структура мозжечка рептилий типична для всех позвоночных: в коре выделяют молекулярный, ганглионарный (слой клеток Пуркинье) и гранулярный слои. Следует отметить, что формирование четкой слоистой структуры коры в мозжечке характерно именно начиная с рептилий, поскольку у амфибий клетки располагаются по всей толще стенки мозжечка. Клетки Пуркинье мозжечка рептилий имеют крупное грушевидное тело (25–35 мкм) и хорошо развитую систему дендритных ветвлений. Клетки Пуркинье контактируют с системой афферентных лиановидных, моховидных и мультислоистых волокон, связывающих мозжечок со спинным мозгом, структурами ствола мозга и другими центрами головного мозга. Также у рептилий начинает формироваться сложная внутренняя межнейронная структура (например, образование гломерулярных структур в гранулярном слое; появление корзинчатых клеток в молекулярном слое).
Эфференты мозжечка образованы аксонами клеток мозжечковых ядер и проходят в ножках мозжечка. По ним информация поступает к ядрам вестибулярного комплекса ствола мозга, красному ядру, нейронам крыши среднего мозга и вышележащим центрам промежуточного (таламус) и конечного мозга. Общим для всех эфферентных трактов мозжечка является то, что через них оказывается влияние мозжечка на моторные центры ствола мозга и спинного мозга.
Средний мозг (Mesencephalon)
В среднем мозге рептилий выделяют крышу среднего мозга (tectum mesencephali, tectum opticum), покрышку (tegmentum) и ножки мозга (pedunculi cerebri). Полостью среднего мозга является так называемый водопровод (aqueductus cerebri). Тектум изучен у рептилий достаточно хорошо и состоит из системы клеточных и волоконных слоев, число которых варьирует в зависимости от вида животных и степени развития зрительной системы. Наиболее сложное строение тектума встречается у хамелеонов, варанов, агам, ряда видов змей (рис. 4).
Основной объем информации тектум получает из сетчатки, причем проекции четко организованы по ретино-топическому принципу: определенные зоны сетчатки проецируются в определенные участки тектума. Развиты связи и с другими отделами головного мозга. Эфференты тектума направляются в вышележащие отделы мозга (в основном в дорсальный таламус), а нисходящие связи направлены в ретикулярную формацию ствола и моторные центры спинного мозга. Последние обеспечивают такие реакции как поворот в сторону добычи/хищника, сочетанные повороты глаз и головы и др. Помимо центров, связанных со зрительным анализатором, у рептилий под тектумом располагается крупная группа нейронов – торус (torus semicircularis), являющийся слуховым центром и гомологом нижних бугорков четверохолмия мозга млекопитающих. В тегментуме среднего мозга располагаются моторные ядра III (глазодвигательного) и IV (блокового) черепно-мозговых нервов, а также крупный центр экстрапирамидной системы мозга – красное ядро (nucleus ruber). Нейроны красного ядра формируют мощные рубро-спинальные и рубро-бульбарные тракты. На границе тегментума и ножек мозга у рептилий находится интерколликулярное ядро, являющееся возможным гомологом черной субстанции (substantia nigra) млекопитающих. Таким образом, средний мозг рептилий содержит нервные центры, связанные со зрительной и слуховой сенсорными системами, а также моторными и премоторными структурами мозга. Все они, в целом, определяют роль среднего мозга, как центра организации и контроля сложных поведенческих реакций рептилий.
Промежуточный мозг (Diencephalon)
Таламус рептилий, как важнейший центр переключения информационных потоков от сенсорных систем в структуры конечного мозга получает значительное развитие. Это выражается в возрастании количества ядерных образований и четко выраженной направленности проекций таламуса в полушария. Основную трудность представляет задача сравнения структур промежуточного мозга рептилий, птиц и млекопитающих и установления их гомологии. Гипоталамус и эпиталамус во многих отношениях оказались структурами более консервативными, сохраняющие сходные черты организации у всех позвоночных. Это, во-первых, связь с эндокринной системой, реализуемые через нейросекреторные центры гипоталамуса и гипофиза. Во-вторых, конвергенция разнообразной информации и участие гипоталамуса в организации поведенческих реакций. В-третьих, связь гипоталамуса со зрительной системой и участие в регуляции биологических ритмов организма.

Рис. 4. Структура тектума среднего мозга змеи Thannophis sirtalis (по: Обухов, Андреева, 2017).
а – эфферентные нейроны тектума, б – интернейроны и эфферентные волокна, 1–6 – эфферентные нейроны: текто-геникулятный (1), текто-истмальный (2), текто-истмо-бульбарный (3), текто-таламический (4), текто-бульбарный прямой (5) и перекрещенный (6), 7–10 – различные типы интернейронов тектума, 11–16 – афферентные волокна из сетчатки (11), тегментума (12), латерального коленчатого тела метаталамуса (13), таламуса (14), перешейка (istmus) (15), моторных ядер среднего мозга (16). SZ-SGP – слои тектума: SZ – stratum zonales, SO – stratum opticum, SPGS – stratum fibrosum et griseum superficiale, SGC – stratum griseum centralis, SAC – stratum album centralis, SGP – stratum griseum periventriculare
Конечный мозг (Telencephalon)
Конечный мозг (Telencephalon) – важнейший отдел ЦНС рептилий. Именно у них формируются формации паллиума, гомологичные корковым структурам мозга птиц и млекопитающих. Паллиум рептилий разделяется на медиальный (архикортекс), дорсальный (возможный гомолог неокортекса) и латеральный (палеокортекс) отделы (рис. 5). Долгое время дорсальный паллиум рассматривали как гомолог неокортекса (слоистая структура, сложные формы нейронов, определенная система связей), однако окончательно этот вопрос не решен. На эту роль претендует и другой отдел полушария: дорсо-вентрикулярный край (ДВК), имеющий паллиальное происхождение, прямые связи с сенсорными ядрами таламуса, сложный нейронный состав. Ряд автором рассматривает ДВК как специфический отдел полушарий мозга рептилий, не имеющий аналога в мозге млекопитающих и развивающийся независимо и параллельно с другими кортикальными формациями других высших позвоночных.

Рис. 5. Структура конечного мозга чешуйчатых рептилий (а), черепах (б) и крокодилов (в), цитоархитектоника дорсо-вентрикулярного края ДВК разного вида и фрагмент нейронной структуры ДВК гадюки (по: Обухов, 1999).
1 – ДВК, 2 – паллиальное утолщение в ДВК, 3 – латеральный мозговой желудочек, 4–6 – разные типы нейронов ДВК, РМm, PMd-m – отделы медиального паллиума, PDd-m, PDd-1 – отделы дорсального паллиума, PL – латеральный паллиум, Nst, Pst – отделы стриатума, Sp – септум, Тol – обонятельный бугорок, na – nucleus accumbens
Субпаллиум рептилий включает собственно стриатум и ряд структур вентромедиальной стенки полушария (обонятельный бугорок, септум, амигдалярный комплекс и ряд мелких ядер), которые имеют сходство с соответствующими структурами мозга высших позвоночных. Таким образом, у рептилий происходит качественный скачок в эволюционном развитии ЦНС. Высшие центры деятельности и регуляции работы всей нервной системы и организма переключаются в структуры промежуточного и конечного мозга. На этом этапе эволюции головного мозга позвоночных закладываются все основные предпосылки к дальнейшему развитию высших отделов мозга, включая корковые формации неокортекса млекопитающих и человека. Поэтому изучение структуры и функционирования головного мозга рептилий имеет ключевое значение в понимании процессов эволюции ЦНС позвоночных животных.